
Aureobasidium Pullulans (Pullaria)
An allergenic mold that can sometimes be found behind wallpaper or on painted or wooden surfaces. Aureobasidium usually develops in a pink, brown or black color. As it ages, it typically turns a darker brown color. The primary health risk of aureobasidium is its ability to cause infections of eye, skin and nails. Because of its potential to cause dermatitis (skin rash), it should never be touched directly with bare skin.
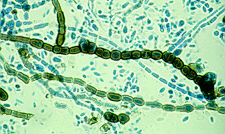 A genus of fungi (family Pseudosaccharomycetaceae) forming yeastlike colonies that are at first dirty white, then streaked with dark green or black, and eventually wholly black and more or less leathery and including a form (P. pullulans) that causes discoloration of pulp and paper. Commonly found on caulk or damp window frams in bathrooms. It is a type of mildew.
A genus of fungi (family Pseudosaccharomycetaceae) forming yeastlike colonies that are at first dirty white, then streaked with dark green or black, and eventually wholly black and more or less leathery and including a form (P. pullulans) that causes discoloration of pulp and paper. Commonly found on caulk or damp window frams in bathrooms. It is a type of mildew.
This yeast-like fungus is commonly found on caulk or damp window frames in bathrooms. Aureobasidium (Pullularia) may be pink or black in color. Although it seldom causes infections, it can be allergenic. This is one type of mold that is a type of mildew. It will grow in cooler climates and along with Cladosporium is commonly found growing on siding.
Pullularia occurs indoors in areas of free water, such as condensate pans, or as a primary colonist of broadloom following a flood. Because its growth form is yeast-like (and are not forcibly discharged), its cells/spores only become airborne through mechanical disruption of contaminated materials or aspiration of contaminated water.
Aureobasidium pullulans is not a primary human pathogen nor is it recognized as a producer of significant mycotoxins. High airborne levels of this fungus have been associated with allergic complaints probably due to respiratory irritation mediated by cell-wall components (e. g. beta glucans, glycoproteins), it has also been known as an irritant, and to cause pulmonary problems (small airway).
Aureobasidium pullulans and the Environment
Polyhydroxy compounds from Aureobasidium pullulans exposed to stress treatments of heat, salt, and simultaneous heat and salt were isolated, identified, and quantified. Results from both thin-layer chromatography (TLC) and high performance liquid chromatography (HPLC) showed that concentrations of trehalose, mannitol, and glycerol increased under stress conditions that induce osmotic- and thermotolerance in A. pullulans. The cellular concentration of trehalose increased in heat-stressed and in simultaneously heat- and salt-stressed cells but not in cells subjected to salt stress alone. Mannitol increased under all stress conditions examined, while an increase in intracellular glycerol was apparent only in salt-stressed cells. The significance of these findings in relation to stress tolerance in salt marsh environments is discussed.
When A. pullulans cells grown at 25 C for 40 h were exposed to sublethal stress conditions (35 C, 4. 5% NaCl, or concurrent treatments of both), the intracellular levels of certain polyhyroxy compounds increased compared to the controls as shown by both TLC and HPLC. Thin-layer chromatograms of cell extracts that were heat shocked at 35 C for 45 min showed a marked increase in trehalose levels compared with controls (data not shown). This represented a near doubling of the mean cell concentration of trehalose compared with untreated controls ( ) as determined by HPLC (compare the results for control cells in with those for heat-shocked cells in).
Although changes in mannitol were not obvious by TLC analysis (due to the overlapping migration of glucose and mannitol and perhaps other sugars in the extracts) HPLC clearly showed a nearly two-fold increase in mannitol as a result of heat shock (). The low level of glycerol in untreated cells did not increase in heat-shocked cells ( ) and glycerol was not detected by TLC. The peak with a retention time of approximately 5 min that appeared in all the HPLC chromatograms appears to be a product of the polysaccharide pullulan which is produced by this fungus. The pullulan “halos” that form around cell pellets during centrifugation produce peaks with a retention time of approximately 5 min when subjected to HPLC.
For treatment, symptoms, and more information see www.mold-survivor.com
Suggested Reading / Abstract
-
- Adler L, Pedersen A, Tunblad-Johansson I., 1982 Polyol accumulation by two filamentous fungi grown at different concentrations of NaCl Physiol Plant 56:139-142
-
- Attfield PV., 1987 Trehalose accumulates in Saccharomyces cerevisiae during exposure to agents that induce heat shock response FEBS Lett 225:259-263 [Medline]
-
- Blomberg A, Adler L., 1993 Tolerance of fungi to NaCl In: Jennings DH, ed. Stress tolerance of fungi. New York: Marcel Dekker. p 209-231
-
- Chatuverdi V, Bartiss A, Wong B., 1997 Expression of bacterial mtlD in Saccharomyces cerevisiae results in mannitol synthesis and protects a glycerol-defective mutant from high-salt and oxidative stress J Bac 179:157-162
-
- Cooke WB., 1959 An ecological history of Aureobasidium pullulans (deBary) Arnaud Mycopathol Mycol Appl 12:1-45
-
- D’Amore T, Crumplen R, Stewart GG., 1991 The involvement of trehalose in yeast stress tolerance J In Microbiol 7:191-196
-
- Hall BG., 1983 Yeast thermotolerance does not require protein synthesis J Bac 156:1363-1365
-
- Henle KJ, Nagle WA, Moss AJ, Herman TS., 1982 Polyhydroxy compounds and thermotolerance: a proposed concatenation Rad Res 92:445-451
-
- Hottiger T, Boller T, Wiemken A., 1987 Rapid changes of heat and desiccation tolerance with changes of trehalose content in Saccharomyces cerevisiae cells subjected to temperature shifts FEBS Lett 220:113-115 [Medline]
-
- Hottiger T, 1989 Correlation of trehalose content and heat resistance in yeast mutants altered in the RAS / adenylate cyclase pathway: Is trehalose a thermoprotectant? FEBS Lett 255:431-434 [Medline]
-
- Hounsa C, Brandt EV, Thevelein J, Hohmann S, Prior BA., 1998 Role of trehalose in survival of Saccharomyces cerevisiae under osmotic stress Microbiology 144:671-680 [Abstract]
-
- Kohlmeyer J, Kohlmeyer E., 1979 Marine mycology: the higher fungi New York: Academic Press. 690 p
-
- Komatsu Y, Kaul SC, Iwahashi H, Obuchi K., 1990 Do heat shock proteins provide protection against freezing? FEMS Microbiol Lett 72:159-162
-
- Lewis JG, Learmonth RP, Watson K., 1995 Induction of heat, freezing and salt tolerance by heat and salt shock in Saccaromyces cerevisiae Microbiology 141:687-694 [Abstract]
-
- Lindquist S., 1986 The heat shock response Annu Rev Biochem 55:1151-1191 [Medline]
-
- Molina FI, Hughes GC., 1982 The Growth of Zalerion maritimum (Linder) Anastasiou in response to variation in salinity and temperature J Exp Mar Biol Ecol 61:147-166
-
- Nass R, Rao R., 1999 The yeast endosomal Na+/H+ exchanger, Hhx 1, confers osmotolerance following acute hypertonic shock Microbiology 145:3221-3228 [Abstract/Full Text]
-
- Panek AC, Vania JJM, Paschoalin MF, Panek D., 1990 Regulation of trehalose metabolism in Saccharomyces cerevisiae during temperature shifts Biochimie 72:77-79 [Medline]
-
- Van Laere A., 1989 Trehalose, reserve and/or stress metabolite? FEMS Microbiol Rev 63:201-210
- Watson K, Dunlop G, Cavicchioli R., 1984 Mitochondrial and cytoplasmic protein synthesis are not required for heat shock acquisition of ethanol and thermotolerance in yeast FEBS Lett 172:299-302 [Medline]




